徕卡显微镜马铃薯块茎的横截面分析
偏振光显微镜(PLM)常常被用在地质学及领域具有显著影响的材料科学中确定复合材料的矿物组成和结构(如纤维)通过使用典型的石英楔补偿器,云母季-waveplates和德Sénarmont赔偿。一个更复杂的组成该设置会带来使用定量极化(通过无论是贝雷或大括号科勒补偿器),在确定个人的各向异性晶体或材料的双折射。可替换地,一个红色-I(λ)板可(耦接与试样厚度的相位差的点的先验知识时)通过消光带和光谱对观察到的比较可以用于确定所发射的光波的相位差的一个米歇尔莱维颜色图表(对于阐明上述结构的双折射)。
尽管如此,使用PLM的目前已下降相对重要性在生物结构的分析和亚细胞器的研究。相反,荧光显微镜为基础的方法已在生物科学取得了由于增加与普遍实现的落射荧光技术在超过阿贝分辨率极限(也称为超分辨率显微镜)的作用的相对重要性,再加上广泛供应和便于标记的利用荧光蛋白质的成像方式。在这简洁(还希望有影响力)的文章中,我们试图评估使用微旋光度(传统的定量PLM的外推)在阐明与分化后卫细胞壁超微结构,而不是淀粉粒形态(从一个全新的上述淀粉颗粒分离出来马铃薯块茎样品)。它进一步希望微旋光作为分析技术的优点将在随后实现的,并进一步通过发展的生物科学和生物成像信息学领域的增强。可以想象,微旋光用荧光显微镜技术的合并将由此产生共鸣以及在这种情况下,通过纳米旋光角[自设计还高度期望的概念用于原位蛋白质组分析的论述来实现潜在的推断,通过一个集成可能实现定量偏光显微镜(qtPLM)和当代(或电流未来增强)超分辨率光学纳米显微镜平台。
材料和方法
徕卡显微镜从头本地化的淀粉颗粒新鲜样品(来自马铃薯块茎的横截面分离)被用作阳性对照,这种评估由于其广泛存在,易于制备[用于透射明场(BF)的PLM技术杂志高双折射和其良好表征的结构。这是对比对的下部(在本文中称为远端)表皮从分枝koenigii分离新鲜制备的载玻片(通过落射PLM观察到的),具有骨架的肌原纤维的商业上制备的幻灯片(具有非常低的双折射通常是悬而未决通过贝雷和去Sénarmont补偿)被用作在这方面(观测来自该滑动收集不包括在本研究中)的阴性对照qtPLM。再加上旋转分析器和一个冷凝器一体型偏振片(用于透射偏振)或对应的入射光偏振器和史密斯:使用HC PL Fluotar50X/0.80 BD物镜(Leica P/N: 566504)进行各向异性识别的标本反射镜立方体(用于落射极化)。作为分析试样进行极化加上判定对象相位差的是通过使用传统的Berek补偿来实现的程度通常是高度双折射的,定量的分析,虽然另一种方法是用于定量目的的相位差(Γobj)中的试样(相对于采用单色光来确定Γobj传统方法)。赔偿德Sénarmont和支撑,科勒的方法是(但)没有用于Γobj或双折射定量在这短短的调查。
结果


A B
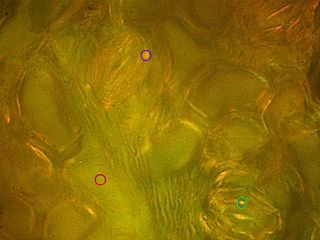
图1:从下diascopic明场极化的马铃薯块茎的图像,而无需使用补偿和B具有相应Berek补偿淀粉粒。双折射的淀粉粒的展览是显而易见的,与Γobj=168.12nm。

A B
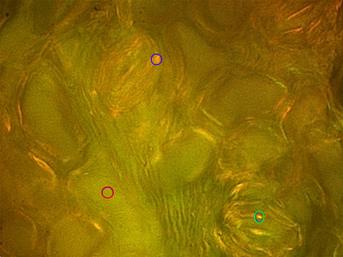
图2:A:M. koenigii下,落射明偏振成像与相应Berek补偿保卫细胞(用夹气孔)。通知它们分别盘旋在蓝色和绿色的保卫细胞的纤维素细胞壁所描绘的,而不是其描绘一黄绿色的色调(Γobj=52.69 nm)的胞质溶胶中的橙色和黄色的多色性(*注:有趣的是,沲移拉曼散射和磷光可能被视为备选假设占该现象,如在后者中,纤维素的特点展示在λ自体荧光与发射Zui大值=420-430nm)。乙:立体图贝雷补偿保卫细胞的PLM成像细胞壁。纤维素微纤丝的致密得多的分布指出,在细胞壁外周,而不是它的中间部分(例如57%/ 40%=1.425倍蓝色圈含有1对保护细胞)。
讨论
结果预期地表明由马铃薯淀粉颗粒和保卫细胞的纤维素细胞壁呈现双折射的差。这两种纤维素和直链淀粉发生(在他们的检测机构)的双折射多糖微纤维;负责相移所描述的方程的一个指定量(Γobj)电子射线相对向O射线,从而影响对所得到的E矢量场的旋度(∇×E)的➀单个的微纤维的空间布置和➁以下(假设不变B)。
考虑将O&E线出现从样品到有相等的幅度,A,与E-和代表分别在y轴和z轴O型轴。那么,我们有[与常数λ和外部补偿(如果有的话)指定为Γcomp]:
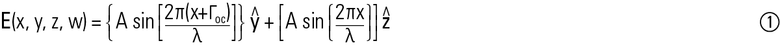
其中x表示横越通过试样和Γoc=Γobj+Γcomp极化波途中的距离。
这必然使得∇×E被导出如下:
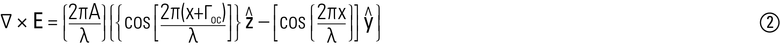
从方程➁以上,则可以观察到∇×E之积变化作为在R3.5的曲线超平面,与Γoc引入一个相位先例翻译到∇×E W-轴。正如所预料的,这在视觉上表现为具有不同Γoc所发送的分析光波的交替放大/消光;从情节∇×E得到随之而来的W-部分将因此允许Γobj的识别哪个暗淡(∇×E)是Zui小的,Γcomp是已知的。然而,如果λ和Γoc是可变的(如通常用于具有不同的双折射的结构时qtPLM下观察时,如淀粉粒和保卫细胞的图1所示的纤维素细胞壁的多色同盟照射标本的情况下和2分别),可以再推测,在λ1..n和Γcompa在Γcomp1..n一些定义λA,暗淡(∇×E)λA是Zui小的Γcompa,占样品/(S观察到的多色性负衰减)(结果)。
此外,通过图2中的各个判定Γobj所观察到的多向色性的分布和定量,主β(1→4)糖苷键的评估保卫细胞的纤维素细胞壁的相对次序可以被推导为平行于气孔孔径的长轴(因为是皮质微管),暗示其在这些细胞中的纤维素合酶GT的结构域被定向在正交的方式相对于较小的气孔轴线。这形成了鲜明的对比不使用微旋光角在确定α(1→4)在图1中的淀粉颗粒糖苷键,对其中的直链淀粉表示含有主要成分的径向取向。
结论
在生物分析的微偏振的一些利用率明显在从20世纪50年代进行了大量的研究 - 1970。然而,偏振镜(作为一个整体元件促进生物研究)已经在过去几十年以实现在荧光显微镜作为生物成像平台更大的兴趣褪在相对重要性,一个概念,通过超分辨率光学显微技术和SIM卡的发展进一步加剧,在其他之中。然而,不同的是需要具体的荧光生物标记物和/或扫描/光场技术(如激光扫描共聚焦显微镜与目前的光纳米显微镜技术的运用),生物PLM提供了一种途径实现的尚未解决的分子结构潜在的异常,但显著的专业知识和工作可能需要创建包含从头双折射值的利益(例如p53和朊病毒[7],主要)的关键蛋白质的数据库。因此diascopic和落射PLM的复苏将是相投在这方面,微旋光确立为传统qtPLM方法外推。